The Growth of Earth’s Supercontinent Spurred Complex Life
 For two billion years, single-celled prokaryotes dominated life on Earth. No one has known just why/when eukaryotes—vastly more complex multi-cellular creatures, like ourselves, with cell nuclei and incredible diversity—arose out of this (comparatively) bland prokaryotic sea.
For two billion years, single-celled prokaryotes dominated life on Earth. No one has known just why/when eukaryotes—vastly more complex multi-cellular creatures, like ourselves, with cell nuclei and incredible diversity—arose out of this (comparatively) bland prokaryotic sea.
But in recent months, hypotheses have been gelling. There has been growing evidence (described in, among other places, the upcoming book The Vital Question) that about 2.5 billion years ago, one prokaryote swallowed another and became its power pack. This dramatically boosted its energy stores, and its ability to rapidly mutate and evolve. The result: a complex creature with organelles and diverse skills: the eukaryote. (See a Bioscience Technology story on a related development here.)
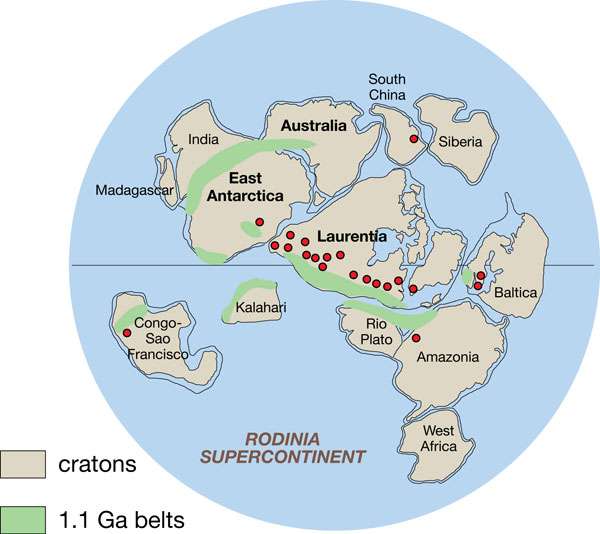
And a (mere) billion years after that, claims a team of Chinese and British scientists in a new Nature Geoscience, the massive supercontinent of Rodinia formed, making oceans churn and shift and lose toxicity on the coasts. This led poisonous hydrogen sulphide to be swept off by tumultuous weather, and uptaken “by extensive continental evaporates on Rodinia,” as the paper puts it.
Ultimately the toxins were replaced, on coasts, by life-giving iron and oxygen, which nurtured to vast numbers “simple but evolved” eukaryotes, Cambridge University lead author Romain Guilbaud, Ph.D., told Bioscience Technology.
Read More: Genesis of Life From Primordial Soup Probed by Scientists
“The interesting new result is that the anoxic Proterozoic ocean stopped having ‘euxinic’ waters—that produce hydrogen sulphide—around its margins roughly a billion years ago,” Tim Lenton, Ph.D., told Bioscience Technology. Lenton, a University of Exeter professor of climate change and earth system science, was not a co-author on the study. But he did much of the research laying groundwork for it.
“The authors infer that this detoxified the waters for eukaryote life forms, for which hydrogen sulphide is toxic, helping facilitate their diversification, and rise to greater ecological prominence around 750 million years ago.”
A crucial period
“In this paper, we have focused on a crucial period of time of Earth's history, in the sense that the Neoproterozoic has witnessed some major biogeochemical revolutions,” Guilbaud, an earth scientist, told Bioscience Technology. “The terminal part of this period looked very different—from an ocean chemistry perspective—from the time before. But the lack of data from this geological interval made any reconstruction of the evolution of the ocean chemistry very difficult.”
The new study let Guilbaud’s group “identify a major chemical (“redox”) transition from a world where, along continental margins where most of the production occurs, toxic, sulphide-rich mid-depth waters were common….to a world where they are almost absent.”
The modern ocean is fully oxygenated, he said, “but at that time, oxygenated waters were restricted to the shallowest parts of the ocean, overlying anoxic waters. This is precisely the nature of these anoxic waters which we determined. Not only did we show this transition, but we also proposed a mechanism which could explain it. Broadly speaking, we believe that the position of the continental masses—all agglomerated as one super-continent around the equator—had an impact on the weathering fluxes to the ocean, in turn impacting the composition of the ocean.”
Coinciding with fossil changes
The retraction of those sulphide-rich zones in the water column had major implications for “the cycle of nutrients—which is tightly linked with redox chemistry—[and] for the ecospace of eukaryotes, as sulphide is a toxin,” said Guilbaud. “So the story got even more exciting when we noticed this chemical transition coincides with a change in the available eukaryotic fossil record. Unfortunately, the fossil record is scarce in the Precambrian, and based on few samples.”
However, Guilbaud said: “This is something to dig into. Perhaps biomarker studies will help in reconciling the fossil and the geochemical record to reconstruct in detail the co-evolution of life and its environment.”
Critiques and comebacks
The paper is not without its critics. Some agree that, despite evidence for subsurface euxinia in mid-Proterozoic marine basins, Neoproterozoic basins generally had subsurface ferruginous (iron-laden) waters. But they note this was an idea first presented by the University of Southern Denmark’s Donald Canfield in a 2008 Science. The evidence from the new paper largely consists of early Neoproterozoic data indicating ferruginous subsurface waters, according to this line of criticism.
Guilbaud agreed the 2008 Canfield paper “illustrated that the later Neoproterozoic subsurface waters were dominantly ferruginous. This is fundamentally different from the picture of the Proterozoic ocean redox structure understood so far, whereby mid-depth, continental margin waters were thought to be euxinic since about 1.8 Ga [billion years ago].”
“However,” Guilbaud told Bioscience Technology, "anoxic, mid-depth data don’t appear before 750 Ma [million years ago], as there was indeed a major gap in the geologic redox record, from about 1.1 Ga to 750 Ma, and what we have done is fill that gap. Our results show the “euxinic wedge” architecture disappeared by about 1 Ga.”
 Another potential problem: iron-speciation data are inherently local, so global statements must be statistical in nature. This requires statistics—and analyses of multiple basins — the Guilbaud paper may not provide. Furthermore, in figure 3, there is an apparent dearth of data in the 1000-800 Ma window, which may be key.
Another potential problem: iron-speciation data are inherently local, so global statements must be statistical in nature. This requires statistics—and analyses of multiple basins — the Guilbaud paper may not provide. Furthermore, in figure 3, there is an apparent dearth of data in the 1000-800 Ma window, which may be key.
Guilbaud responded that, “indeed, the iron speciation does not give a global perspective, but it is not local either. This gives us a broad local, or best regional, idea. It is also the only calibrated method to identify several redox states of the water column. It is therefore the most robust redox proxy used by redox geochemists. In order to have a more global picture, we do need to analyze multiple basins and work on statistics. We did both. We analyzed five different basins from different settings throughout the margins of supercontinent Rodinia, and its epicontinental seas.”
He continued: “A problem, which applies to any of us, is the scarcity in the provision of well-preserved geological rocks which cover the time period of interest, and our samples were beautifully preserved. Yes, there are not many data between about 1 Ga and about 850 Ma. But before our study, there were none. In addition to our analyses, we compiled about 1000 data points from anoxic basins which potentially could bear euxinic waters, in order to get a robust statistical context for your results.”
Lenton agreed. “The authors concentrate on ocean margins where the—limited—evidence for eukaryotes exists. Sampling of deep time is always sparse, so building up a global picture will always be difficult. I think all they are saying is, from the inevitably limited sampling, earlier ocean margins commonly show evidence for euxinia, and then that goes away in the early Neoproterozoic.”
It has also been noted that work from many groups, including that of Yale University biogeochemist Timothy Lyons, have showed that, even before the Neoproterozoic, subsurface water masses were predominantly ferruginous. So there may be limited reason to believe the loss of euxinic water masses would have caused global biological upheaval.
Guilbaud responded that indeed, “many groups, including Tim Lyons’, have worked on the mid-Proterozoic, and depicting a ‘global ocean’ redox architecture.” (One such Lyons study is titled: “Widespread iron-rich conditions in the mid-Proterozoic ocean.”) “These studies have helped draw the redox structure of the ocean since about 1.8 Ga, as I was mentioning earlier.”
However, Guilbaud told Bioscience Technology: “What we showed is different: the continental, mid-depth settings stopped being euxinic. So we are saying the nature of anoxia has changed within the productive margins, where biology thrives. This is what we focused on here.”
Said Lenton: “Everyone agrees the deep ocean was predominantly ferruginous before the Neoproterozoic. Simon Poulton was the key person who showed that. [But] that is not the point the present paper is addressing. It is talking about whether there was a ‘wedge’ of euxinic water at shallow-intermediate depths in upwelling margins/shelf seas, which could have had evolutionary consequences. The data analysed from China probably lies in the 1000-800 Ma window. The problem is a broader one of getting reliable estimates of age on the Neoproterozoic record.”
There has also been some criticism of the group’s idea that the decline of sulphidic water masses removes an environmental barrier to eukaryotic evolution. While it makes sense, it may not necessarily be correct for above reasons—and because the timing of the major eukaryotic diversification may not match the team’s proposed environmental change-over.
Referring to his earlier comments, Guilbaud added: “There have been many major eukaryotic diversification episodes, and we are discussing one of them. The timing is always tricky, as rocks are dated with a large error, and the geochemical timescale diverges significantly from the biological timescale, making it difficult to correlate time-wise the fossil record from the geochemical one. However, in the case of our study, one can observe a trend, whereby there is a conspicuous change in the fossil record where we also observe the disappearance of euxinic conditions in settings where biological production occurs.”
Lenton said he agreed timing may not match. “I too think there is quite a potential mismatch in timing, between environmental change and rise of eukaryotes, but this is back to the broader problem of dating the Neoproterozoic record. We need more reliable age estimates, and an overall stratigraphy for the Proterozoic. Currently, unless we have the geochemistry and the fossils/biomarkers in the same section, then temporal causality is difficult to infer. Still, there are other exciting locations in China where early eukaryote fossils have been discovered. So it will be interesting to examine the geochemical signature of those sections.
No reservations
The bottom line, Lenton told Bioscience Technology, is that he has “no reservations about the geochemical analyses, nor the fact that hydrogen sulphide is toxic to eukaryotes at sufficient concentration.” Still, he said: “I can think of other mechanisms that could have helped shift the ocean from partially euxinic to wholly ‘ferruginous’—anoxic and iron rich—conditions – in particular, the interactions between ocean redox state and the cycling of the crucial limiting-nutrient, phosphorus.”
So a critical next step, Lenton said, may be to “test the hypothesis that supercontinent formation was key to the shift in ocean chemistry, by modelling the cycles of iron, sulphur, oxygen, carbon, phosphorus and nitrogen, and the feedbacks between them. The suggested mechanism should make some testable predictions for the effects on nutrient and carbon cycling.”
He concluded that “a broader challenge is to test what, if anything, in the environment held back eukaryote evolution? The jury is out on whether eukaryote evolution was just inherently slow, or whether it really was environmentally constrained—by hydrogen sulphide, lack of oxygen, or something we haven’t thought of yet.”