Effects of Supercontinent Formation on Complex Life: Part II
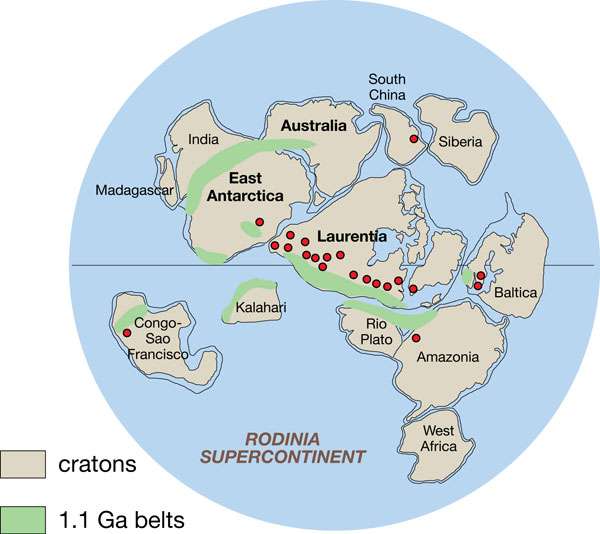 Recently, Chinese and British scientists reported in Nature Geoscience that the formation of earth supercontinent Rodinia, 1 billion years ago, may have swept toxic hydrogen sulphide away from Proterozoic ocean coasts. This significantly accelerated the growing complexity of eukaryotic life there, the team surmised, as hydrogen-sulphide was replaced by eukaryote-friendly iron.
Recently, Chinese and British scientists reported in Nature Geoscience that the formation of earth supercontinent Rodinia, 1 billion years ago, may have swept toxic hydrogen sulphide away from Proterozoic ocean coasts. This significantly accelerated the growing complexity of eukaryotic life there, the team surmised, as hydrogen-sulphide was replaced by eukaryote-friendly iron.
Not everyone agrees. In early June, Bioscience Technology ran the first of two articles featuring back-and-forth between scientists on this fascinating topic.
This week, Bioscience Technology offers the thoughts of Cambridge University lead author, and earth scientist, Romain Guilbaud, Ph.D., and University of California, Riverside, geochemist Timothy Lyons, Ph.D.
Loss of widespread sulphide as key
Lyons, who has written much on what he calls “widespread iron-rich conditions” of the mid-Proterozoic ocean, noted to Bioscience Technology the Guilbaud team argues that “expanded eukaryotic diversification in the early Neoproterozoic was driven by water chemistry on ocean margins that shifted to being more dominantly anoxic and iron-rich (ferruginous), rather than anoxic and hydrogen-sulphide-containing (euxinic). Euxinia was relatively common during the preceding time interval. This is certainly a novel idea, and quite different from the most popular hypothesis that Neoproterozoic diversification was instead a product of more broadly and consistently oxygenated conditions in the ocean, including persistently higher levels of oxygen in the surface ocean. My colleagues and I have argued in favor of oxygenation — although that oxygen-dependent eukaryotic diversification, based on fossil and geochemical records, is more easily placed at ca. 0.8 to 0.75 billion years ago, rather than the authors’ ‘event’ at ca. 1.0 Ga [billion years ago].”
In response, Guilbaud told Bioscience Technology his team does embrace the idea that “the distribution and the diversification of eukaryotes is tightly linked with oxygen levels in the surface ocean. After all, eukaryotic forms which are preserved in the fossil record, and most definitely in the assemblages described in our contribution, were likely aerobes and thrived in oxygenated, shallow waters.
"There is  biomarker evidence for a major biological change around 0.75 billion years ago, whereby eukaryotes began to dominate the biology. However, we have identified that a conspicuous change within eukaryotic assemblages broadly coincides with a change in the nature of mid-depth anoxia, around 1 Ga.”
biomarker evidence for a major biological change around 0.75 billion years ago, whereby eukaryotes began to dominate the biology. However, we have identified that a conspicuous change within eukaryotic assemblages broadly coincides with a change in the nature of mid-depth anoxia, around 1 Ga.”
Lyons said that Guilbaud’s team, using iron data, suggested in their paper that “the loss of widespread sulphide is the key. This certainly is a clever idea, but there are issues to continue pondering. For example, their model assumes that oxygen is not the controlling factor, but rather sulphide toxicity was the throttle. In other words, their conceptual model envisions replacing earlier widespread sulphidic (euxinic) waters with anoxic (iron-rich, ferruginous) waters, and it is the decrease in sulphide (H2S) at mid-water depths that favored eukaryotes in the shallower oxic waters. Specifically, less sulphide in deeper waters meant less upward mixing of toxic, eukaryote-stifling H2S into the surface waters, which, they claim, favored eukaryotic diversification in those waters.”
Lyons further questioned whether “upward mixing of anoxic waters might have been able to have a similar impact on delaying favorable conditions for eukaryotes. I’m struggling to imagine eukaryotes taking off in an evolutionary way because episodically sulphidic waters were replaced in time by episodically anoxic waters.”
Guilbaud responded that eukaryotes have designed various strategies to survive episodes of anoxia. “During anoxic incursions into surface waters, iron oxidation occurs at a rate which is several orders of magnitude higher than sulphide oxidation,” he told Bioscience Technology. In other words, when iron-rich waters mix upward with oxic surface waters, “iron precipitates as particles at the chemocline (the zone which separates oxic from oxygen-free waters). By contrast, when H2S-rich waters meet the chemocline, sulphide can persist in the shallow waters. Sulphide is a toxin to all eukaryotes, whereas iron is also a nutrient, and we argue that the cessation of sulphide stress onto eukaryote forms did open up new opportunities to diversify.”
Conclusions that ‘swing for the fences’
Lyons noted however that, even though the authors of the Nature Geoscience paper “have anticipated this challenge, I am convinced that the extremely low organic contents in the China samples, which are central to their story, contributed to (or controlled) the absence of sulphidic conditions at those sites. Organic matter is required by microbes to drive reduction of sulphate to sulphide. So a lack of that essential ingredient could explain why sulphide was missing at a particular location, while potentially still being abundant elsewhere in the world ocean in more organic-rich settings. In other words, I remain concerned that local controls might be the story here, which would mute their assertion about global controls on eukaryotic diversification. Since their conclusions swing for the fences, it would be more convincing to see similar conditions in other parts of the ancient ocean over the same time interval. Independent that verification, the results are ambiguous.”
Guilbaud countered that organic matter is indeed an essential ingredient for sulphate reduction, “as it is also for the bacterial reduction of iron. When produced, sulphide reacts with iron and cannot build up in the water column unless the provision in iron is overwhelmed by the production of sulphide.”
Therefore, Guilbaud said, his team argued that “organic matter contents control the presence of euxinia only if the highly reactive iron supply has been exhausted. Organic contents in the China sections are indeed extremely low. They are, however, much higher (and in sufficient amounts to support sulphate reduction) in the Amundsen, the Officer, the Amadeus, and the Spitzbergen basins, which all point towards a retraction of euxinia in the time period studied. We agree that we always need more data to strengthen (or disprove) a story. It seems that for this time period, however, local exceptions may concern settings with high organic carbon and sulphide-rich conditions rather than the opposite.”
Lyons said that if the Guilbaud team's estimates were correct, the amount of sulphate in the ocean “was still 1 to 3 mM [millimolars], which is far beyond the expected amount of iron dissolved in seawater at that time, and should have been enough to support sulphide production and accumulation. The absence of sulphide, then, becomes more likely a product of low organic content locally. In fact, their curve shows estimates for earlier sulphate concentrations at or below these Neoproterozoic predictions—during mid-Proterozoic time periods well known for relatively widespread euxinia (sulphidic waters). Their response would be that the Neoproterozoic was also marked by greater iron input through continental weathering, which buffered against the buildup of hydrogen sulphide. But this is really hard to test.”
Guilbaud responded that sulphate inputs to the ocean “mainly derive from the dissolution of sulphide minerals (pyrite) on the continent. Through rivers, this process brings sulphate, but also iron to the ocean, in which pyrite can form again and remove back both iron and sulphate.”
The crucial point, Guilbaud said, is that “it is not only the amount of dissolved iron which plays a role, but the totality of the so-called ‘highly reactive iron’ minerals, which will all react with sulphide and prevent it from accumulating in the water column. These minerals cover the majority of the total Fe [iron] riverine discharge. And at a time where ocean sulphate contents were about 10 fold smaller than today—this is a maximum estimate—the balance between sulphate and iron becomes very sensitive to smaller input changes.”
Massive burial of gypsum on Rodinia
Finally, Lyons said, the authors of the Nature Geoscience paper “imagine that a/the trigger for this hypothesized global decline in sulphide accumulation on the ocean margins was decreasing sulphate delivery to the oceans. And they have attributed this declining input to massive burial of sulphate evaporates (gypsum) on the newly assembled Rodinia supercontinent. If they are correct, the key here is that sulphate was trapped on the continents, and therefore not delivered to the oceans, making it harder to establish sulphidic conditions in the ocean.”
If, on the other hand, any of those evaporite basins were connected to the ocean, Lyons noted, “the ‘sudden’ appearance of abundant gypsum deposition would mean the opposite—that is, higher sulphate in the ocean, so that evaporation could lead to supersaturation and precipitation. And that higher sulphate would have meant a greater likelihood of euxinia given sufficient local organic matter availability to support abundant microbial sulfate reduction. Earlier seawater struggled to form gypsum, even when evaporated, because sulphate was very low, yet there was euxinia. I don’t know these gypsum-containing deposits and basins well enough to comment further on the question of a seawater contribution to those sites, but it would be a critical question to ask the authors and an area of important further study.”
Concluded Guilbaud: “The issue here is the paucity of evaporitic data for the time interval studies, and we were careful not to over-interpret the geologic record. However, other studies do infer enhanced sulphate-containing deposits on large inner-seas, which support our proposed mechanism here.”